EL SISTEMA DE COMPLEMENTO
ANTECEDENTES HISTÓRICOS DEL SIS. DE COMPLEMENTO

Ya antes del fin del siglo XIX Ehrlich había usado el término "complemento" para designar la actividad del suero que podía complementar la capacidad de los anticuerpos específicos de lisar bacterias. Pero es Jules Bordet quien descubre (1895) este componente, caracterizado frente a los anticuerpos por su termolabilidad. En 1907 Ferrata comienza a caracterizar algunos de sus componentes recurriendo a métodos de diálisis. Por motivos meramente cronológicos, los componentes iban recibiendo denominaciones a base de números tras la letra "C" conforme se iban descubriendo. Por esta razón, su orden de actuación no guarda en general relación con su nomenclatura.
¿Qué es el sistema del complemento?
Como se sabe, la inmunidad innata cumple un papel de erradicación de los agentes patógenos gracias a procesos humorales y celulares. En el caso de la inmunidad humoral no específica, se puede mencionar el sistema del complemento.

Ante la entrada de un patógeno se ponen en marcha diversas vías capaces de erradicarlo, cada agente generará la activación de distintos sistemas. En el caso de la inmunidad inespecífica (innata) humoral, uno de sus protagonistas es el Sistema del complemento. Sin embargo, este protagonismo se pierde por ejemplo con la infección viral, puesto que los interferones alfa y beta cobran un papel fundamental.
En sí, el sistema del complemento es un conjunto de proteínas generadas en su mayoría por células hepáticas que se activan mediante clivaje, de manera ordenada, consecutiva, conformando una cascada de reacciones que culminan con la entrada masiva de iones y agua a la célula. Tal efecto provoca la muerte celular por explosión. Su accionar es pro inflamatorio, por tal motivo se encuentra estrictamente regulado.
| |
| |
|
Las tres rutas comparten las últimas fases, consistentes en el ensamblaje, sobre la superficie del microorganismo, del denominado complejo de ataque a la membrana.
ACTIVACIÓN DE LA VÍA CLÁSICA
La vía clásica comienza casi siempre estimulada por la formación de complejos antígeno-anticuerpo. Ya se ha visto que tanto la Ig G como la Ig M pueden activar al complemento (a excepción de Ig G de tipo 4). Esta vía puede separarse en dos estadíos, uno primario en el que intervienen C1 - C2 - C3- C4 y una segunda etapa en común con el resto de las vías en la que intervienen C5 - C6 - C7 - C8 y C9.
Primer etapa:

Figura 1 - Activación del Sistema del Complemento
1 - Se forma el complejo antígeno-anticuerpo. La interacción entre estos suscita cambios en la conformación molecular del fragmento Fc de la inmunoglobulina en cuestión. Estos cambios generan un sitio de adhesión para C1 expuestos en la sección CH2 de la partícula del anticuerpo.
2 - La molécula de C1 está formada por subunidades C1q (que posee seis brazos helicoidales y un tallo), 2 subunidades C1r y 2 subunidades C1s. Todas estas subunidades se encuentran estabilizadas por el ión calcio.
3 - Se desarrolla la interacción complejo anticuerpo-antígeno + cabezas globulares de la estructura helicoidal de C1q. Se requiere, por lo menos, la participación de dos sitios de unión a C1q. La Ig G solo posee un sitio de adhesión por partícula, por tal motivo se requieren como mínimo dos moléculas Ig G.
La Ig M por su parte, expone más sitios de adhesión cuando se encuentra en forma de 'grapa'. Esto explica el
porqué la Ig M es más propensa a activar el sistema.
4 - La interacción entre la región Fc y C1q activa a C1r adquiriendo propiedades de serinproteasa, C1r . C1r por su parte, clivará a C1s generando su forma activa: C1s.
5 - C1s tiene dos sustratos, C2 y C4. Será primero C4 quien interactúe con él produciendo C4a (pequeño) y C4b (grande). C4b se fijará a la membrana de la célula blanco, C4a difunde hacia el plasma. C2 por su parte, encuentra un sitio de unión en C4b, como todo este complejo se encuentra cerca de C1s , C2 es clivado por ésta generando C2a y C2b. En este caso, el fragmento más pequeño es C2b (que difunde hacia el plasma) quedando unido a C4b el fragmento C2a.
6 - Queda formado el complejo C4b2a (C4b + C2a). A éste se le denomina "Convertasa de C3", porque justamente C3 es su sustrato a quien activa generando C3a y C3b (tener en cuenta los pequeños fragmentos generados C3a, C4a, C5a para más adelante, se les llama anafilatoxinas). La convertasa C4b2a es capaz de generar decenas de moléculas de C3b, por ello se le suele llamar a este paso "amplificador".
7- C3b se unirá a C4b2a generando C4b2a3b, complejo que recibe el nombre de "Convertasa de C5". La subunidad C3b de este complejo se unirá a C5 para que el resto lo hidrolize generando C5a y C5b. C5b es un componente clave para la formación del "sistema de ataque" a la membrana celular. Cabe destacar que no toda el C3b generado participa en el complemento, también hay una fracción que difunde hacia el plasma con función de opsonización.
Segunda Etapa:
En esta etapa se da la convergencia de las tres vías. Participan aquí C5b, C6, C7, C8 y C9 con la finalidad de formar una estructura molecular conocida como complejo de ataque a la membrana. Éste se insertará en la misma a fin de permitir el ingreso masivo de iones, partículas y agua provocando la pérdida de la estabilidad celular y su muerte.
1 - C5b se une a la membrana celular, siempre en la región hidrófila externa de esta.
2 - C5b se une a C6 formando C5b6.
3 - C5b6 interactúa con C7 formando un complejo que comienza a exponer regiones hidrófobas capaces de penetrar a la sección interna de la bicapa lipídica, el complejo recibe el nombre de C5b67
4 - C8 se une a C5b67 formando un pequeño poro de 10 amstrong capaz de destruir glóbulos rojos pero no células con núcleo.
5 - Finalmente, varias unidades C9 se unen al complejo C5b678 generando un poro que mide entre 70 y 100 Amstrong. Por él puede ingresar a la célula una gran cantidad de iones, moléculas y agua a la vez que se pierden electrolitos importantes desembocando en la muerte de la misma


ACTIVACIÓN DE LA VÍA ALTERNATIVA
La ruta alternativa se activa directamente sobre la superficie de muchos microorganismos. Opera varios días antes de que entre en acción la ruta clásica (la clásica tiene que esperar a que se hayan producido anticuerpos).

Activación "al ralentí" o "marcapasos"
En el suero, en una situación normal (en ausencia de infección) se está produciendo continuamente una activación limitada que produce sólo pequeñas cantidades de C3b*:
El enlace tioéster interno del C3 se hidroliza espontáneamente en agua, dando una forma activada llamada C3i. Esto es lo que se conoce como activación al ralentí (activación tick-over).
El C3i actúa ahora como sitio de unión para el factor B, generando el complejo C3iB, sobre el que actúa el factor D, que rompe el B unido para generar Ba y el complejo C3iBb, que actúa como una C-3 convertasa en fase fluida. Como tal, escinde el C3 en C3a y C3b*.
Pero como este C3b* está en fase fluida, la mayor parte de él se hidroliza por agua y se inactiva. Ahora bien, si por casualidad alguna molécula de C3b* se topa con una superficie no propia (p. ej., la membrana de una bacteria), se une covalentemente a ella e inicia el bucle de amplificación de la ruta alternativa.
ACTIVACION POR LA VÍA DE LECTINAS

La ruta comienza por la acción de la proteína de unión a mananos (MBP). Se trata de un componente parecido estructuralmente al C1q: hexámeros con 18 cadenas polipeptídicas idénticas enrolladas de tres en tres. Los hexámeros de MBP se pueden unir con dos unidades de C1r y dos de C1s, pero parece que va acompañada de su propia serín-proteasa (denominada MASP), que muestra casi 40% de homología con C1r o C1s.
La MBP se une preferentemente a los extremos de manosa, fucosa y glucosamina de polisacáridos o glucoproteínas de membrana de gran variedad de bacterias. De modo similar a lo que ocurre con el complejo C1, cuando la MBP se engarza con esos carbohidratos, sufre un cambio conformacional que a su vez activa a su serín-proteasa (MASP). Una vez activada, la MASP actúa secuencialmente sobre C4 y C2, para producir una C3-convertasa de la ruta clásica.
LAS FUNCIONES DEL SISTEMA DEL
COMPLEMENTO
A. Lisis de células

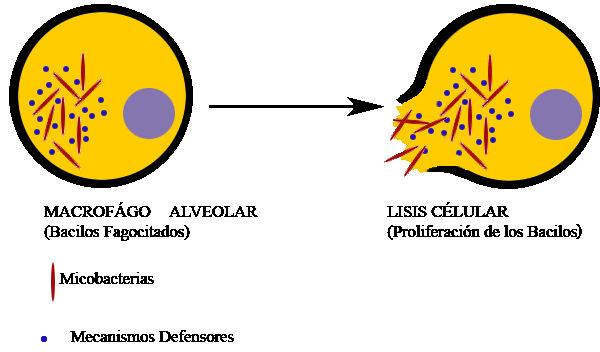
El MAC (membrane attack
complex/complejo de ataque a la membrana) puede lisar bacterias gram-negativas,
Parásitos, virus encapsulados, eritrocitos y células nucleadas.
Las bacterias gram-positivas son
bastante resistentes a la acción del complemento.
B. Respuesta inflamatoria
 Los pequeños fragmentos que
resultan de la fragmentación de componentes del complemento, C3a, C4a y C5a,
son llamados anafilotoxinas. Estas se unen a receptores en células cebadas y
basófilos. La interacción induce su degranulación, liberando histamina y otras
sustancias farmacológicamente activas. Estas sustancias aumentan la permeabilidad
y vasodilatacion.
Los pequeños fragmentos que
resultan de la fragmentación de componentes del complemento, C3a, C4a y C5a,
son llamados anafilotoxinas. Estas se unen a receptores en células cebadas y
basófilos. La interacción induce su degranulación, liberando histamina y otras
sustancias farmacológicamente activas. Estas sustancias aumentan la permeabilidad
y vasodilatacion.
Así mismo, C3a, C5a y C5b67 inducen monocitos
y neutrófilos a adherirse al endotelio para iniciar su extravasación.
 C. Opsonización
C. Opsonización

C3b es la opsonina principal del
complemento. Los antígenos recubiertos con C3b se unen a receptores específicos
en células fagocíticas, y así la fagocitosis es facilitada.
 D. La neutralización de virus
D. La neutralización de virus

C3b induce la agregación de
partículas virales formando una capa gruesa que bloquea la fijación de los
virus a la célula hospedera. Este agregado puede ser fagocitado mediante la
interacción de receptores del complemento y C3b en células fagocíticas.

E. Eliminación de complejos inmunes

Los complejos inmunes (complejos
antígeno-anticuerpo circulantes) pueden ser eliminados de la circulación si el complejo
se une a C3b. Los eritrocitos tienen receptores del complemento que interactúan
con los complejos inmunes cubiertos por C3b y los lleva al hígado y al bazo
para su destrucción
BIBLIOGRAFÍA
https://www.ugr.es/~eianez/inmuno/cap_16.htm
inmunologia.eu/sistemas-y-procesos/sistema-del-complemento
http://resumendeinmunologia.blogspot.com/2011/07/via-clasica-del-sistema-del-complemento.html
VÍDEOS EDUCATIVOS
VIDEO ELABORADO POR LA MESA 3
INTEGRANTES DE LA MESA 3
Carla Fernanda Arevalo Basulto
Azucena Abigali Fomperosa Juarez
-Emily Damaris Guzman Palacios
-Nayeli Sabet Mora Alvarado
-Andrik Roberto Reyes Dominguez
-Julian Ramirez Azamar
-Trinidad Seba Chigo
Azucena Abigali Fomperosa Juarez
-Emily Damaris Guzman Palacios
-Nayeli Sabet Mora Alvarado
-Andrik Roberto Reyes Dominguez
-Julian Ramirez Azamar
-Trinidad Seba Chigo
REFLEXIÓN SOBRE EL USO DE LAS TIC'S
Gracias a las tecnologías de la información y de la comunicación logramos alimentar nuestros conocimientos empleando la información que necesitamos para obtener trabajos de calidad como lo es un blog educativo que sirve para integrar de diversas fuentes conceptos para el aprendizaje.
Buuuuu, que feo .
ResponderEliminarMentira, está muy bien .:)
Muy buena información, qué buen blog.
ResponderEliminarMuy buena información :v
ResponderEliminarMuy buena información está bien buena.
ResponderEliminarMuy buena información está bien buena.
ResponderEliminarExcelente blog y muy buena información, le quedó muy bonito 👍🏻
ResponderEliminarMuy buena información, muy completa y el diseño llamativo
ResponderEliminarMuy bien su resumen, los felicito.
ResponderEliminarMi nombre es Lucy y escribo desde Kansas City, Estados Unidos. Quiero usar su plataforma para compartir la buena obra del Dr. Jekawo, cuyo sitio web es www.drjekawo.com y su correo electrónico es drjekawo@gmail.com. Es un verdadero médico herbolario que prepara medicinas a base de hierbas que curaron por completo el herpes y el VPH de mi hija. Estoy tan agradecida que le prometí al Dr. Jekawo que compartiría su buen trabajo con cualquier persona que necesite mejorar su salud o curarse de alguna enfermedad. El Dr. Jekawo cura todas las enfermedades e infecciones en hombres y mujeres, como el VIH, el virus del herpes, la diabetes, el cáncer, las enfermedades respiratorias, las enfermedades del sistema inmunológico y el VPH. El Dr. Jekawo es un hombre enviado por Dios para sanar a los enfermos, al igual que el Dr. Sebi.
ResponderEliminar